MITOSIS

Mitosis is
the process, in the cell cycle, by which a cell duplicates into two genetically
identical daughter cells. In mitosis, chromosomes in the cell
nucleus are separated into two identical sets of chromosomes, each in its
own nucleus. In general, mitosis is followed immediately by cytokinesis, which
divides the cytoplasm,organelles, and cell membrane, and later karyokinesis,
which divides the nucleus, dividing cell into two containing roughly equal
shares of these cellular components. Mitosis and cytokinesis together
define themitotic (M) phase of the cell cycle—the division of the
mother cell into two daughter cells, genetically identical to each other and to
their parent cell. This accounts for approximately 20% of the cell cycle.Mitosis occurs only
in eukaryotic cells and the process varies in different
groups. For example, animal sundergo an "open" mitosis,
where the nuclear envelope breaks down before the chromosomes
separate, while fungi such as Asperqillus nidulans and yeast undergo a "closed" mitosis,
where chromosomes divide within an intact cell
nucleus. Prokaryotic cells, which lack a nucleus, divide by a process
called binary fission.The process of mitosis is fast and highly complex.
The sequence of events is divided into stages corresponding to the completion
of one set of activities and the start of the next. These stages are
prophase,prometaphase, metaphase, anaphase, and telophase. During mitosis, the
pairs of chromatids condense and attach to fibers that pull the sister
chromatids to opposite sides of the cell. The cell then divides in
cytokinesis, to produce two daughter cells.
Because cytokinesis often occurs in conjunction with mitosis, "mitosis" is often used interchangeably with "mitotic phase". However, there are many cells where mitosis and cytokinesis occur separately, forming single cells with multiple nuclei. The most notable occurrence of this is among the fungi and slime molds, but is found in various groups. Even in animals, cytokinesis and mitosis may occur independently, for instance during certain stages of fruit fly embryonic development. Errors in mitosis can either kill a cell through apoptosis or cause mutations. Certain types of cancer can arise from such mutations.
Because cytokinesis often occurs in conjunction with mitosis, "mitosis" is often used interchangeably with "mitotic phase". However, there are many cells where mitosis and cytokinesis occur separately, forming single cells with multiple nuclei. The most notable occurrence of this is among the fungi and slime molds, but is found in various groups. Even in animals, cytokinesis and mitosis may occur independently, for instance during certain stages of fruit fly embryonic development. Errors in mitosis can either kill a cell through apoptosis or cause mutations. Certain types of cancer can arise from such mutations.
INTERPHASE
The mitotic
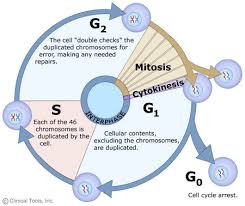
phase is a relatively short period of the cell cycle. It alternates with
the much longer interphase, where the cell prepares itself for the
process of cell division. Interphase is divided into three phases: G1 (first
gap), S
(synthesis), and G2 (second
gap). During all three phases, the cell grows by
producing proteins and cytoplasmic organelles. However, chromosomes are
replicated only during the S
phase. Thus, a cell grows (G1), continues to grow as it duplicates its
chromosomes (S), grows more and prepares for mitosis (G2), and finally it divides (M) before restarting
the cycle. All these
phases in the cell cycle are highly regulated, mainly via proteins. The phases
follow one another in strict order and there are "checkpoints" that
give the cell the cues to proceed from one phase to another. There is also a
fourth section in Interphase where the cell has the option to enter G0. Cells continue on through this cell cycle until
they become too crowded; at that point they will exit the cell cycle and enter G0. This reaction is called contact inhibition or density-dependent
inhibition. Altogether interphase takes up roughly
90% of a cell's lifespan.
PROPHASE
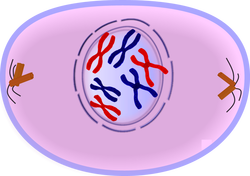
During this stage the cell
prepares to divide by dissolving the membrane around the nucleus and the
chromatin condenses into chromosomes. Normally, the genetic material in the
nucleus is in a loosely bundled coil called chromatin.
At the onset of prophase, chromatin fibers become tightly coiled, condensing
into discrete chromosomes. It is
crucial for the reader to note that chromatin is a complex consisting of both
DNA and specific proteins. Since the genetic material has already been
duplicated earlier in S phase, the replicated chromosomes have two sister
chromatids, bound together at the centromere by the cohesin protein complex. Chromosomes are
typically visible at high magnification through a light microscope.
Also inside the nucleus, the nucleolus in the nucleus disappears from view. This is noteworthy because the cell does not need to divide the nucleolus right away. It will later reform when the nucleus divides completely.
Close to the nucleus are structures called centrosomes, consisting of a pair of centrioles, and actin, a halo of microtubule fragments. Centrioles are found in most eukaryotic animal cells. The centrosome is the coordinating center for the cell's microtubules. A cell inherits a single centrosome at cell division, which is replicated by the cell with the help of the nucleus before a new mitosis begins, giving a pair of centrosomes. The two centrosomes nucleate microtubules (which may be thought of as cellular ropes or poles) to form the spindle by polymerizing soluble tubulin. Molecular motor proteins then push the centrosomes along these microtubules to opposite sides of the cell. Although centrioles help organize microtubule assembly, they are not essential for the formation of the spindle, since they are absent from plants, and centrosomes are not always used in mitosis.
Also inside the nucleus, the nucleolus in the nucleus disappears from view. This is noteworthy because the cell does not need to divide the nucleolus right away. It will later reform when the nucleus divides completely.
Close to the nucleus are structures called centrosomes, consisting of a pair of centrioles, and actin, a halo of microtubule fragments. Centrioles are found in most eukaryotic animal cells. The centrosome is the coordinating center for the cell's microtubules. A cell inherits a single centrosome at cell division, which is replicated by the cell with the help of the nucleus before a new mitosis begins, giving a pair of centrosomes. The two centrosomes nucleate microtubules (which may be thought of as cellular ropes or poles) to form the spindle by polymerizing soluble tubulin. Molecular motor proteins then push the centrosomes along these microtubules to opposite sides of the cell. Although centrioles help organize microtubule assembly, they are not essential for the formation of the spindle, since they are absent from plants, and centrosomes are not always used in mitosis.
METAPHASE
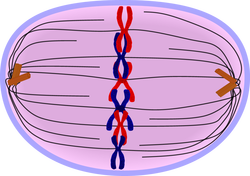
After the microtubules have found and attached to the kinetochores in prometaphase, the two centrosomes start pulling the
chromosomes through their attached centromeres towards the two ends of the
cell. As a result, the chromosomes come under longitudinal tension from the two
ends of the cell. The centromeres of the chromosomes, in some sense, convene
along the metaphase plate or equatorial
plane, an imaginary line that is right in between the two centrosome poles. This line is called the spindle
equator. This even alignment is due to the counterbalance of the pulling powers
generated by the opposing kinetochores, analogous to a tug-of-war between
people of equal strength. In certain types of cells, chromosomes do not line up
at the metaphase plate and instead move back and forth between the poles
randomly, only roughly lining up along the midline.
Because proper chromosome separation requires that every kinetochore be attached to a bundle of microtubules (spindle fibres), it is thought that unattached kinetochores generate a signal to prevent premature progression to anaphase without all chromosomes being aligned. The signal creates the mitotic spindle checkpoint.
Because proper chromosome separation requires that every kinetochore be attached to a bundle of microtubules (spindle fibres), it is thought that unattached kinetochores generate a signal to prevent premature progression to anaphase without all chromosomes being aligned. The signal creates the mitotic spindle checkpoint.
ANAPHASE
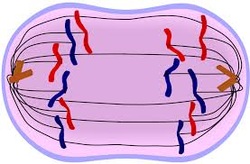
When every kinetochore is
attached to a cluster of microtubules and the chromosomes have lined up along
the metaphase plate, the cell proceeds to anaphase.Two events then occur:
First, the proteins that bind sister chromatids together are cleaved. These sister
chromatids now become separate daughter chromosomes, and are pulled apart by
shortening kinetochore microtubules and move toward the respective centrosomes
to which they are attached. The cleaved centromeres go first while the
chromatids trail behind. They all look as if they are trying to grab at their
partners, because they become shaped like a V.
Next, the polar microtubules elongate, pulling the centrosomes (and the set of chromosomes to which they are attached) apart to opposite ends of the cell. The force that causes the centrosomes to move toward the ends of the cell is still unknown, although there is a theory that suggests that the rapid assembly and breakdown of microtubules may cause this movement. At the end of anaphase the kinetochore microtubules all degrade.
Next, the polar microtubules elongate, pulling the centrosomes (and the set of chromosomes to which they are attached) apart to opposite ends of the cell. The force that causes the centrosomes to move toward the ends of the cell is still unknown, although there is a theory that suggests that the rapid assembly and breakdown of microtubules may cause this movement. At the end of anaphase the kinetochore microtubules all degrade.
TELOPHASE
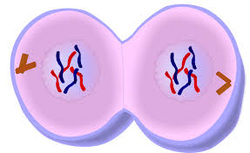
Telophase is a reversal of
prophase and prometaphase events. It "cleans up" the after effects of
mitosis. At telophase, the polar microtubules continue to lengthen, elongating the cell even more.
Corresponding daughter chromosomes attach at opposite ends of the cell. A new
nuclear membrane, using the membrane vesicles of the parent cell's old nuclear
membrane, forms around each set of separated daughter chromosomes (though the
membrane does not enclose the centrosomes) and the nucleolus reappears. Both
sets of chromosomes, now surrounded by new nuclei, begin to "relax"
or decondense back into chromatin. Mitosis is complete, but cell division is
not.
MEIOSIS I
Meiosis is
the process by which replicated chromosomes undergo two nuclear divisions to
produce four haploid cells, also called meiocytes (sperms and eggs). Diploid (2n) organisms rely on meiosis to
produce meiocytes, which have half the ploidy of the parents, for sexual
reproduction. Halving the ploidy in meiocytes is essential for restoring the
genetic content of the zygote to that of the parents. Meiosis uses
similar mechanisms as those employed during mitosis to accomplish the
separation and redistribution of chromosomes. However, several features,
namely, the pairing and genetic recombination between homologous chromosomos, are unique
to meiosis.
The steps leading up to meiosis are similar to those of mitosis – the centrioles and chromosomes are replicated. The amount of DNA in the cell has doubled, and the ploidy of the cell remains the same as before, at 2n. In meiosis I, the phases are analogous to mitosis: prophase I, metaphase I, anaphase I, and telophase I (below figure). Meiosis I proceeds directly to meiosis II without going through interphase.
Meiosis I is unique in that genetic diversity is generated through crossing over and random positioning of homologous chromosomes (bivalent chromosomes). In addition, in meiosis I, the chromosomal number is reduced from diploid (2n) to haploid (n) during this process. (See figure below, where meiosis I begins with a diploid (2n = 4) cell and ends with two haploid (n = 2) cells.) In humans (2n = 46), who have 23 pairs of chromosomes, the number of chromosomes is reduced by half at the end of meiosis I (n = 23).
The steps leading up to meiosis are similar to those of mitosis – the centrioles and chromosomes are replicated. The amount of DNA in the cell has doubled, and the ploidy of the cell remains the same as before, at 2n. In meiosis I, the phases are analogous to mitosis: prophase I, metaphase I, anaphase I, and telophase I (below figure). Meiosis I proceeds directly to meiosis II without going through interphase.
Meiosis I is unique in that genetic diversity is generated through crossing over and random positioning of homologous chromosomes (bivalent chromosomes). In addition, in meiosis I, the chromosomal number is reduced from diploid (2n) to haploid (n) during this process. (See figure below, where meiosis I begins with a diploid (2n = 4) cell and ends with two haploid (n = 2) cells.) In humans (2n = 46), who have 23 pairs of chromosomes, the number of chromosomes is reduced by half at the end of meiosis I (n = 23).
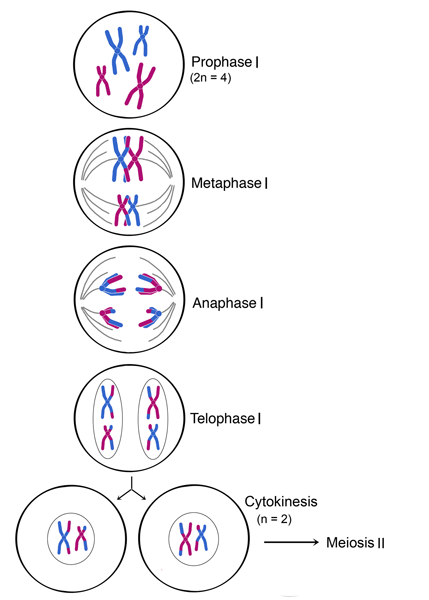
PROPHASE I
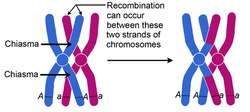
During prophase I, chromosomal condensation allows chromosomes to be viewed under the microscope. In late prophase I, homologous chromosomes (also called bivalent chromosomes, or bivalents) pair laterally, or side-by-side. At this time they are said to be in synapsis. During synapsis, crossovers – cross-connections that form from breakage and rejoining between sister chromatids – can occur between the paired bivalents, leading to genetic recombination (exchange of genetic material) between the strands involved. The point where a crossover occurs is called a chiasma (plural chiasmata) (see below figure). In figure below, following crossing over, the blue and red chromosomes, which originally carried AA and aa alleles, respectively, now carry Aa alleles in both chromosomes at the end of prophase I. Note that these bivalents have two chromosomes and four chromatids, with one chromosome originating from each parent.
Düzenlemek için buraya tıklayın.
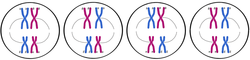
In metaphase I, each pair of bivalents (two chromosomes, four chromatids total) align on the metaphase plate. This is different from metaphase in mitosis, where all chromosomes align single file on the metaphase plate. The position of each chromosome in the bivalents is random - either parental homolog can appear on each side. This means that there is a 50-50 chance for the daughter cells to get either the mother's or father's homolog for each chromosome (see figure below). As shown in the below figure, during metaphase I, bivalents from either parent can align on either side of the cell. In an organism with two sets of chromosomes, there are four ways in which the chromosomes can be arranged, resulting in differences in chromosomal distribution in daughter cells after meiosis I. (A diploid organism with 2n chromosomes will have 2n possible combinations or ways of arranging its chromosomes during metaphase I.)
ANAPHASE I
In anaphase I, homologous chromosomes separate. Homologous chromosomes, each containing two chromatids, move to separate poles. Unlike in mitosis, the centromeres do not split and sister chromatids remain paired in anaphase I.
TELOPHASE I AND CYTOKINESIS
In telophase I, the homologs of each bivalent arrive at opposite poles of the cell, and a new nuclear membrane forms around each set of chromosomes. Cytokinesis then divides the cell into two daughter cells. Each of the two daughter cells is now haploid (n), with half the number of chromosomes per nucleus as in meiosis I. In some species, the nuclear membrane briefly forms around the chromosomes, while in others it does not. The cell now proceeds into meiosis II, with the chromosomes remaining condensed.
MEIOSIS II
Chromosomal replication does not occur between meiosis I and meiosis II; meiosis I proceeds directly to meiosis II without going through interphase. The second part of the meiosis, meiosis II, resembles mitosis more than meiosis I. Chromosomal numbers, which have already been reduced to haploid (n) by the end of meiosis I, remain unchanged after this division. In meiosis II, the phases are, again, analogous to mitosis: prophase II, metaphase II, anaphase II, and telophase II (see figure below). As shown in the figure below, meiosis II begins with two haploid (n = 2) cells and ends with four haploid (n = 2) cells. Notice that these four meiocytes are genetically different from one another. In humans (2n = 46), who have 23 pairs of chromosomes, the number of chromosomes remains unchanged from the beginning till the end of meiosis II (n = 23).
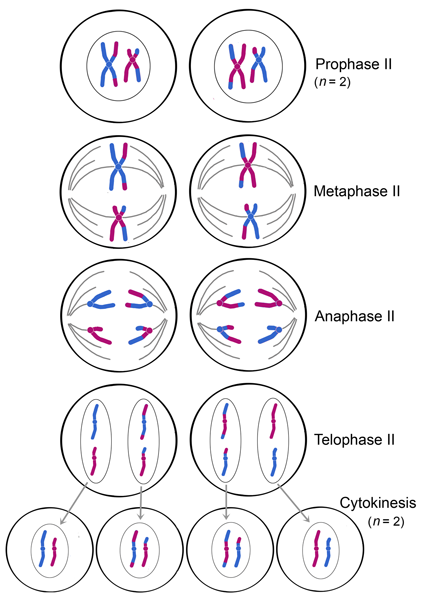
PROPHASE II
Spindle fibers reform and attach to centromeres in prophase II.
METAPHASE II
The chromosomes align on the metaphase plate during metaphase II in preparation for centromeres to divide in the next phase.
ANAPHASE II
In anaphase II, chromosomes divide at the centromeres (like in mitosis) and the resulting chromosomes, each with one chromatid, move toward opposite poles of the cell.
TELOPHASE II AND CYTOKINESIS
Four haploid nuclei (containing chromosomes with single chromatids) are formed in telophase II. Division of the cytoplasm during cytokinesis results in four haploid cells. Note that these four cells are not identical, as random arrangements of bivalents and crossing over in meiosis I leads to different genetic composition of these cells.
In humans, meiosis produces genetically different haploid daughter cells, each with 23 chromosomes that consist of one chromatid. These haploid cells become unfertilized eggs in females and sperm in males. The genetic differences ensure siblings of the same parents are never entirely genetically identical.
In humans, meiosis produces genetically different haploid daughter cells, each with 23 chromosomes that consist of one chromatid. These haploid cells become unfertilized eggs in females and sperm in males. The genetic differences ensure siblings of the same parents are never entirely genetically identical.